ホーム
研究内容
メンバー
研究業績
学生募集
アクセス・連絡先
写真集
English
English
研究内容
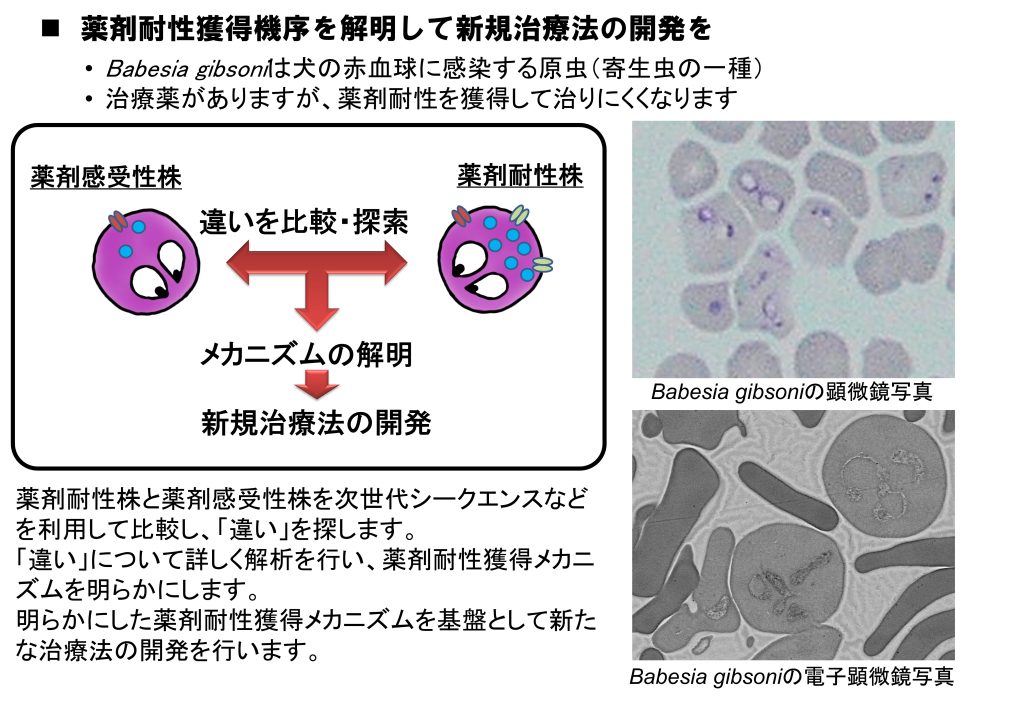
1. 犬バベシア症の制圧にむけて
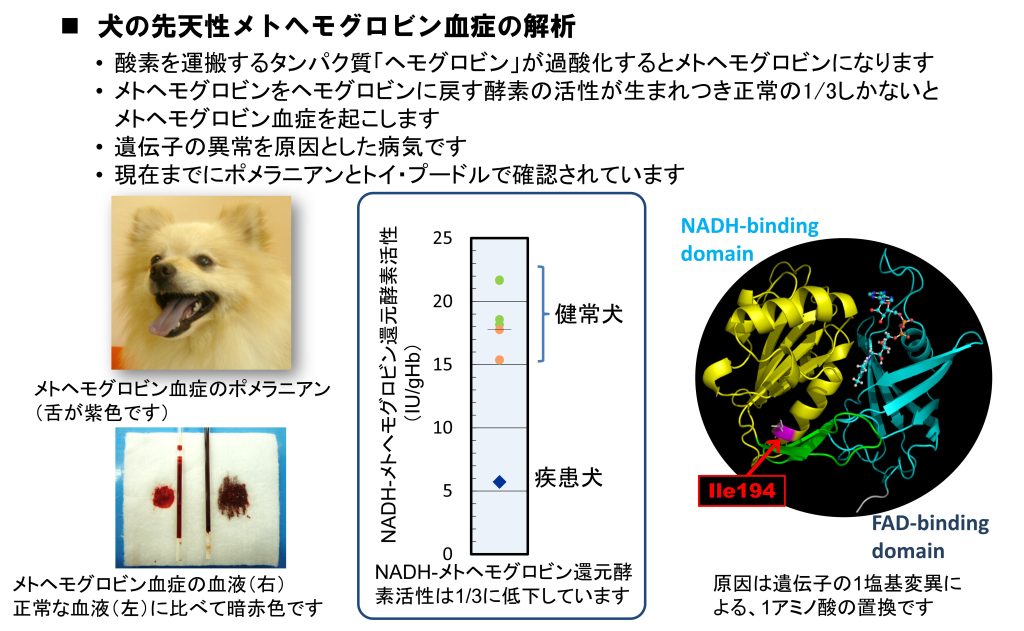
2. 犬猫の遺伝病に関する解析
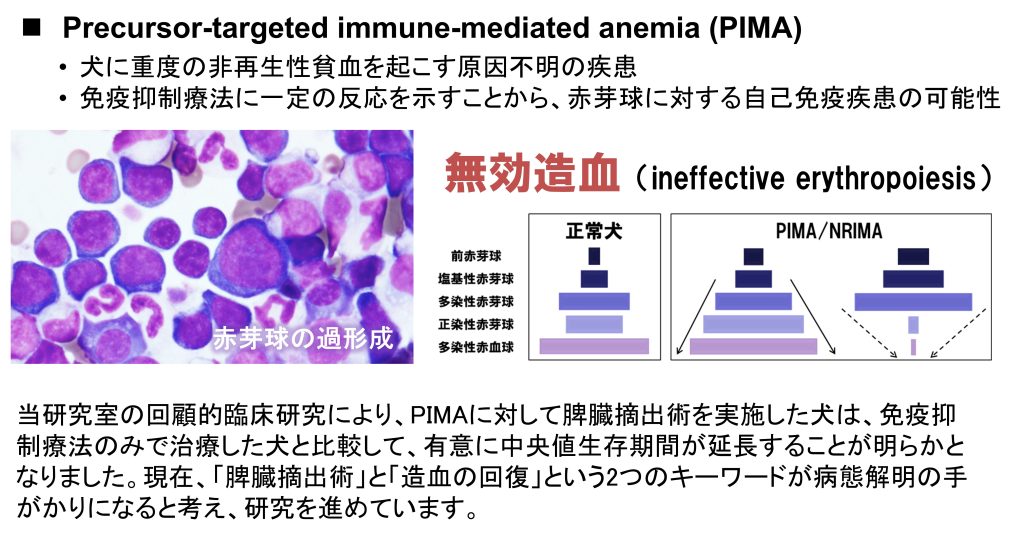
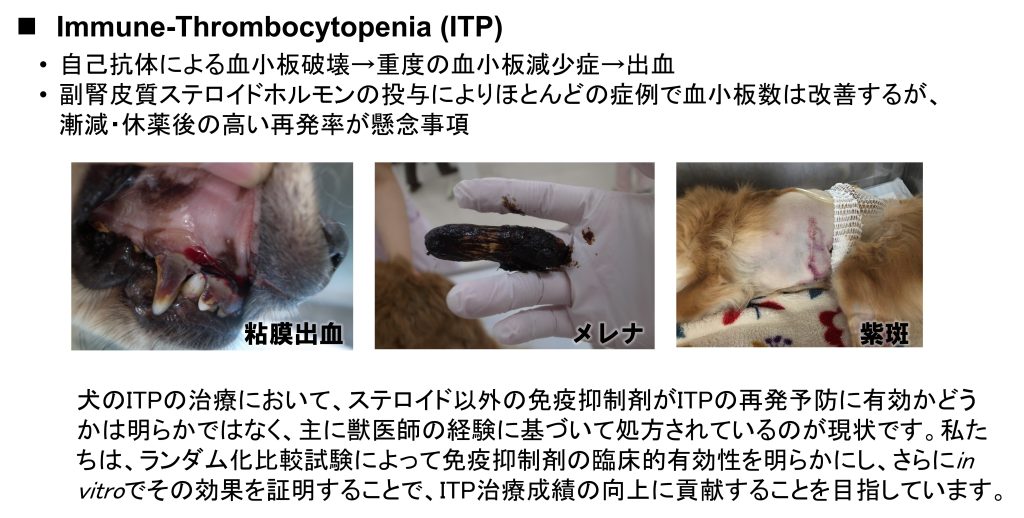
3. 犬の免疫介在性血球減少症の治療成績向上に挑む